نتورینگ کسب و کارها و استارتاپ ها در GlobalTechHarbor
 by
by در گوشهای از دنیای پرتلاطم فناوری، گروهی با همت و ذوق، زیر آسمان نوآوریها به وجود آمدهاند. ما، گروه “GlobalTechHarbor”، نه تنها جایگاهی برای فعالیتهای …
بازی ، سینما ، تکنولوژی ، مد ، سبک زندگی ، آشپزی و گردشگری
در گوشهای از دنیای پرتلاطم فناوری، گروهی با همت و ذوق، زیر آسمان نوآوریها به وجود آمدهاند. ما، گروه “GlobalTechHarbor”، نه تنها جایگاهی برای فعالیتهای …
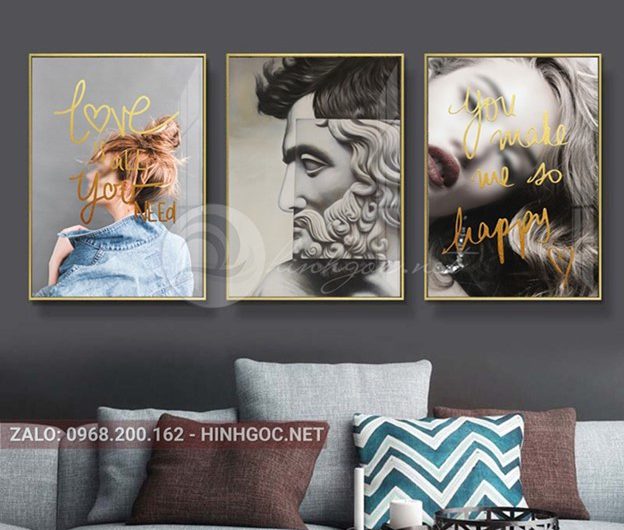
خرید تابلو عکس دکوراتیو یکی از مهمترین عناصر در دکوراسیون داخلی است که به عنوان یک اثر هنری میتواند تأثیر بسزایی بر زیبایی و ارزش …
معیارهای مهم در انتخاب بهترین گیم سرور چیست؟ اگر بخواهیم بدانیم که بهترین معیارها برای انتخاب سرور گیم چیست، باید موارد متعددی را مدنظر قرار دهیم. درادامه، …
منبع مطلب زیر سایت tgju مقاله سایز لوله مسی کولر گازی می باشد. سایز لوله مسی کولر گازی شما چه مقدار است؟ آیا از اندازه …
مک دارو یک داروخانه اینترنتی است که به مشتریان خود امکان خرید داروها را به صورت آنلاین فراهم میکند. این سرویس جدید و مبتکرانه به …
انتخاب بهترین بیمه تکمیلی: وقتی صحبت از بهترین بیمه تکمیلی به میان میآید، یعنی باید اطلاعات دقیقی در خصوص انواع بیمههای درمان تکمیلی بدست آورد. در اکثر …
ردیاب و دزدگیر خودرو با برند سام ترک (samtrack) یک ردیاب یا GPS چند منظوره با محدوده کنترل عریض است، که معمولا طیف وسیعی از …
موم یا وکس زنبور عسل، یکی از مواد طبیعی جهت حفاظت و سلامت پوست محسوب میشود. از موم به عنوان یک امولسیفایر در ساخت انواع …
این لباسهای فشردهکننده با طراحیهای متنوع و ویژگیهای منحصر به فرد، برای افرادی که به دنبال تغییر در ظاهر و افزایش اعتماد به نفس هستند، …
وقتی صحبت از بهترین بیمه تکمیلی به میان میآید، یعنی باید اطلاعات دقیقی در خصوص انواع بیمههای درمان تکمیلی بدست آورد. در اکثر مواقع، عوامل تاثیر …